December 16th, 2014
Key Findings
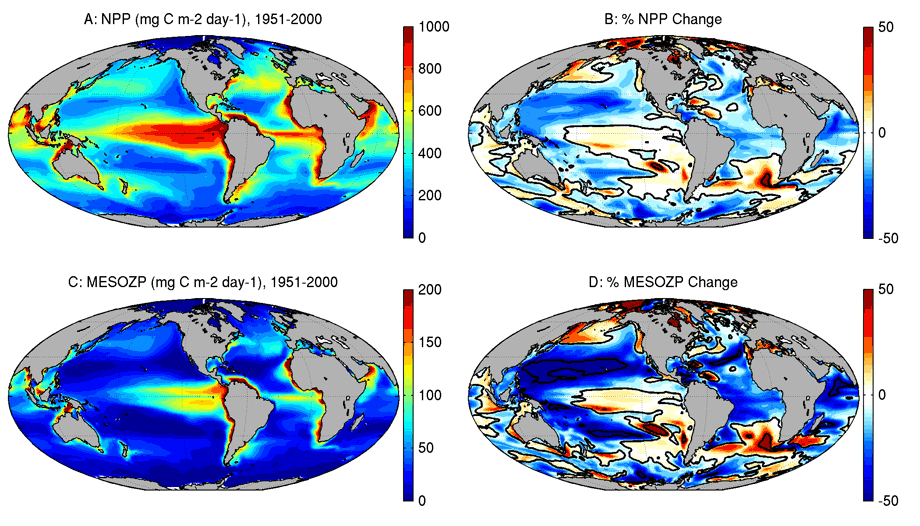
- GFDL’s Earth System Model projects that changes in the production of large zooplankton may be twice as large as those for phytoplankton (Figure 1) by the end of this century.
- There is potential for much larger regional zooplankton productivity changes than global average trends, even greater than 50% in some cases.
- Such changes raise the prospect for significant regional redistributions of marine resources.
- The “amplification” of ocean productivity changes at higher trophic levels arises from changes in consumer-prey interactions that control the flow of energy from phytoplankton to fish.
Charles A. Stock, John P. Dunne, Jasmin G. John. Biogeosciences. DOI: 10.5194/bg-11-7125-2014.
Summary
Earth System Models (ESMs) project that climate change will lead to approximately 1-10% declines in global ocean phytoplankton productivity by the end of the 21st century, under high carbon emissions scenarios. This decline results from projected increases in ocean stratification under global warming, which hinders the supply of deep ocean nutrients to the well-lit surface ocean.
In this paper, we show that climate change projections with GFDL’s Earth System Model further suggest that changes in the production of large zooplankton (krill, for example) may be twice as large as those for phytoplankton (Figure 1). The results also highlight the potential for much larger regional zooplankton productivity changes than global average trends, even greater than 50% in some cases. Such changes raise the prospect for significant regional redistributions of marine resources with implications for regional economies and food security.
We demonstrate that the “amplification” of ocean productivity changes at higher trophic levels arises from changes in the planktonic food web, or the set of consumer-prey interactions that control the flow of energy from phytoplankton to fish. First and foremost, declining phytoplankton production doesn’t just decrease the amount of food zooplankton eat. It also decreases the fraction of that food that contributes to production. Zooplankton require energy to simply maintain their activities before any new biomass is produced. This “maintenance” requirement increases as the ocean warms. With less phytoplankton to consume and increasing maintenance, the fraction of food available for production also declines, accentuating changes in phytoplankton productivity for zooplankton.
Second, large phytoplankton are hurt most by higher stratification. Large cells make for less effective scavenging for the diminished nutrient supply expected under climate change, relative to smaller-celled counterparts. While large zooplankton such as krill can eat large phytoplankton directly, small phytoplankton are too small and must first be consumed by smaller zooplankton before benefitting large zooplankton. Less than 30% of the energy in small phytoplankton, however, remains once the small zooplankton’s energetic needs are accounted for. Suppression of large phytoplankton under climate change thus also decreases the efficiency of energy transfer from phytoplankton to fish, amplifying percent productivity declines for higher trophic levels relative to phytoplankton.
The combined impact of the two mechanisms above is felt most strongly in the already unproductive sub-tropical ocean gyres – the large “blue” regions north and south of the equator in panel A of the Figure. In these regions, organisms are already surviving with small energy surpluses relative to demands, and diminished phytoplankton production is sharply felt.
Trophic amplification is not all bad. The two mechanisms work in reverse, amplifying productivity increases, most notably in many regions of the Arctic. Also, enhanced ocean stratification reduces interruptions in ocean productivity caused by deep winter mixing, leading to a tighter coupling between phytoplankton prey and zooplankton consumers. These positive effects, however, are outweighed by the prevalence of phytoplankton productivity declines and the amplification of those declines by the mechanisms above.
There is considerable work to be done to refine projections of ocean productivity changes in this paper, particularly in understanding the complex patterns of regional change in Figure 1. While different ESMs agree on many large-scale patterns, they often disagree regionally. These differences need to be reconciled to fully understand implications for regional fisheries yields. Also, while the planktonic food web dynamics in GFDL’s marine ecosystem model have been closely vetted, some aspects of the model are poorly constrained by existing observations and laboratory work. We are working with NOAA and academic colleagues to fill these gaps.